Објави:



Типови синдикалних ћелија и њихове карактеристике
Тхе спојеви ћелија они су контактни мостови који постоје између цитоплазматских мембрана између суседних ћелија или између ћелије и матрице. Зглобови зависе од типа испитиваног ткива, наглашавајући везе између епителних, мишићних и нервних ћелија.
У ћелијама постоје молекули везани за адхезију између њих. Међутим, потребни су додатни елементи који повећавају стабилност везивања ткива. Ово се постиже са ћелијским спојевима.
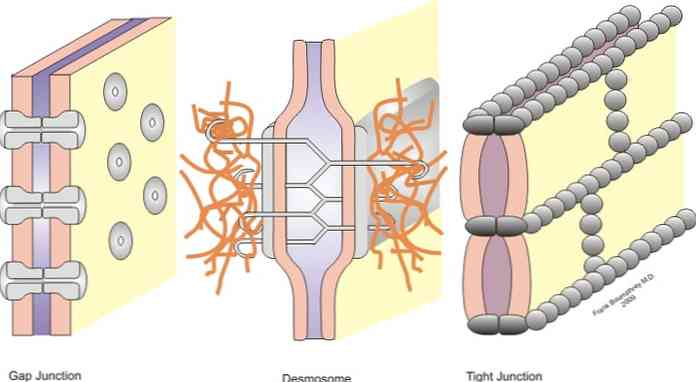
Спојеви су класификовани у симетричне зглобове (уски зглобови, десмосоми у појасу и резни зглобови) и асиметрични зглобови (хемидесмосоми)..
Уски спојеви, десмосоми појаса, десмосоми и хемидесмозоми су зглобови који омогућавају сидрење; док се спојеви у прорезу понашају као мостови уједињења између суседних ћелија, дозвољавајући размену растворених супстанци између цитоплазми.
Кретање раствора, воде и јона се одвија кроз и између појединачних ћелијских компоненти. Тако постоји трансцелуларни пут који се контролише низом проводника и транспортера. За разлику од парацелуларног пута, који је регулисан контактима између ћелија - то јест, ћелијски спојеви.
У биљкама налазимо целуларне спојеве који личе на крижања, названи плазмодезми. Иако се разликују у својој структури, функција је иста.
Са медицинске тачке гледишта, одређени недостаци у ћелијским спојевима доводе до стечених или насљедних болести узрокованих оштећењем епителне баријере.
Индек
- 1 Карактеристике
- 2 Типови
- 2.1 - Уске везе
- 2.2 - Спојнице прореза или зазора
- 2.3 - Сидрење или везивање
- 2.4 -Хемидесмосомас
- 3 Ћелијски спојеви у биљкама
- 4 Медицинска перспектива
- 5 Референце
Феатурес
Живи организми су састављени од дискретних и разноврсних структура које се називају ћелије. Оне су ограничене плазматском мембраном која их чува одвојено од екстрацелуларне средине.
Међутим, иако су састојци живих бића, они не личе на цигле, јер нису изоловани једни од других.
Ћелије су елементи који се међусобно комуницирају и са изванстаничним окружењем. Стога, мора постојати начин да ћелије формирају ткива и комуницирају, док мембрана остаје нетакнута.
Овај проблем је решен захваљујући присуству ћелијских спојева који постоје у епителу. Ови спојеви се формирају између две суседне ћелије класификоване према функцији сваког у симетричним и асиметричним зглобовима..
Асиметричне везе припадају хемидесмосомима, а на симетричним везама блиске везе, десмосом на појасу, десмосоми и прорезе. Затим ћемо детаљно описати сваки од синдиката.
Типови
-Узани спојеви

Уски спојеви, такође познати у литератури као оклузивна чворишта, су сектори у ћелијским мембранама суседних ћелија које су интимно повезане - као што име "уско спајање" означава.
Под просечним условима, ћелије су раздвојене на удаљености од 10 до 20 нм. Међутим, у случају уских спојева, ова удаљеност је значајно смањена и мембране обе ћелије су дотакнуте или чак спојене.
Типичан уски спој се налази између бочних зидова сусједних ћелија на минималној удаљености од њихових апикалних површина.
У епителном ткиву све ћелије формирају синдикате овог типа како би остали уједињени. У овој интеракцији, ћелије се налазе у облику узорка који подсјећа на прстен. Ови синдикати покривају читав периметар.
Протеини који су укључени у уске везе
Оцлудина и Цлаудина
Подручја блиског контакта окружују целу површину ћелије. Ове регије формирају контактне траке анастомозе трансмембранских протеина познатих као оклудин и клаудин. Термин анастомоза односи се на унију одређених анатомских елемената.
Ова два протеина припадају групи тетраеспанина. Карактерише се са четири трансмембранска домена, две спољашње петље и два релативно кратка цитоплазматска репа.
Уочено је да оклудин интерагује са четири друга молекула протеина, који се називају оклудин зонуле и скраћено ЗО. Ова последња група обухвата протеине ЗО 1, ЗО 2, ЗО 3 и афуна.
Цлаудин је, с друге стране, породица од 16 протеина који чине низ линеарних фибрила у уским спојевима, што омогућава овом синдикату да преузме улогу "баријере" у парацелуларном путу..
Нецтинас и ЈАМ
Нектини и адхезиони молекули синдиката (скраћено акронимом на енглеском језику ЈАМ), такође се појављују у уским спојевима. Ове две молекуле се налазе као хомодимери у интрацелуларном простору.
Нектини су повезани са актин филаментима помоћу афадинског протеина. Ово последње изгледа да је од виталног значаја, јер у делецијама гена који кодира за афадин код глодара, они доводе до смрти ембриона..
Функције уских спојева
Ови типови веза између ћелија и ћелија обављају две основне функције. Први је да се одреди поларитет ћелија у епителу, одвајајући апикалну домену од базолатерала и спречавајући непотребну дифузију липида, протеина и других биомолекула..
Као што смо споменули у дефиницији, ћелије епитела су груписане у прстен. Ова структура одваја апикалну површину ћелије од латералних и базалних, што успоставља диференцијацију између домена.
Ово одвајање се сматра једним од најважнијих концепата у проучавању физиологије епитела.
Друго, уски спојеви спречавају слободан пролаз супстанци кроз слој епителних ћелија, што се преводи у баријеру парацелуларног пута..
-Спојнице или прорези

Раскиди или расцјепи се налазе у подручјима без ограничења цитоплазматске мембране између сусједних станица. У расцепљеном споју, цитоплазме ћелија су повезане и настаје физичка веза где се може појавити пролаз малих молекула..
Ова класа зглобова се налази у готово свим епителима, као иу другим типовима ткива, где служе веома разноврсне сврхе..
На пример, у неколико ткива, прорезе могу да се отварају или затварају као одговор на ванћелијске сигнале, као што је случај са неуротрансмитерима допамином. Присуство овог молекула смањује комуникацију између класа неурона у ретини, као одговор на повећани интензитет светлости.
Протеини који су укључени у зглобове цепања
Прорезе су формиране од протеина који се називају конексини. Дакле, "цонекон" се добија уједињењем шест конексин мономена. Ова структура је шупљи цилиндар који пролази кроз цитоплазматску мембрану.
Конексони су распоређени тако да се између цитоплазми сусједних ћелија ствара канал. Поред тога, конексони имају тенденцију да се агрегирају и формирају неку врсту плоча.
Функције разрезаних спојева
Захваљујући формирању ових веза, може доћи до кретања одређених молекула између суседних ћелија. Величина молекула који се транспортује је одлучујућа, оптимални пречник је 1.2, као што су калцијумови јони и циклични аденозин монофосфат.
Конкретно, неоргански јони и молекули растворљиви у води могу да се пренесу из ћелијске цитоплазме у сусједну цитоплазму.
Концентрације калцијума играју кључну улогу у овом каналу. Када се концентрација калцијума повећа, аксијални канали имају тенденцију да се затварају.
На тај начин, шупљи спојеви активно учествују у процесу електричног и хемијског повезивања између ћелија, као што је то случај у ћелијама срчаног мишића, које су одговорне за пренос електричних импулса.
-Сидрење или везивање

Испод уских спојева налазе се сидрени спојеви. Генерално, они се налазе у близини апикалне површине епитела. У овој групи можемо разликовати три главне групе, зонула адхеренс или десмосома у појасу, макула адхеренс или десмосом точка и десмосом \ т.
У овом типу спојева, суседне ћелијске мембране које су спојене зонулама и адхеренским макулама одвојене су ћелијском растојањем које је релативно широко - ако их упоредимо са минималним простором који постоји у случају уских спојева..
Међустанични простор заузимају протеини који припадају породици кадхерина, десмоглеина и десмоколина везаних за цитоплазматске плоче које имају друге протеине назване десмоплакуина, плацоглобина и плацофилина.
Класификација спојева за сидрење
Зонула адхеренс
Као иу случају уских спојева, у сидреним спојевима посматрамо и облик распореда у облику прстена или ремена. Зонула адхеренс је повезана са актиним микрофилмом, кроз интеракцију два протеина: кадхерина и катенина.
Мацула адхеренс
У неким случајевима, ова структура је позната као десмосома, то је тачкаста веза која је повезана са средњим филаментима формираним од кератина. У том контексту, наведене структуре кератина се називају "тонофилиманетос". Филаменти се протежу од једне тачке до друге у епителним ћелијама.
Поинт десмосомас
Оне обезбеђују снагу и ригидност епителним ћелијама. Стога се сматра да је његова главна функција повезана са јачањем и стабилизацијом суседних ћелија.
Десмосоми се могу упоредити са врстом заковица или заварених спојева, јер личе на одвојене ситне тачке, а не на континуалне траке.
Овај тип зглобова налазимо у интеркалираним дисковима који повезују кардиоците у срчаном мишићу и менингама које покривају спољну површину мозга и кичмене мождине..
-Хемидесмосомес

Хемидесмосоми спадају у категорију асиметричних спојева. Ове структуре имају функцију сидрења базалног домена епителне ћелије са базалном ламином.
Термин хемидесмосом се користи зато што ова структура изгледа, буквално, "средњи" десмосом. Међутим, са становишта њихове биохемијске композиције, оба синдиката су потпуно другачија.
Важно је појаснити да су десмосоми одговорни за спајање суседне ћелије са другом, док је функција хемидесмосома да се придружи ћелији са базалном ламином..
За разлику од макуле или десмосома, хемидесмосоми имају другачију структуру, која се састоји од: цитоплазматске ламеле повезане са средњим филаментима и плоче спољашњих мембрана, која је одговорна за спајање хемидесмосома са базалном ламином, помоћу анцхор филамент.
Једна од функција хемидесмосома је да повећа укупну стабилност епителних ткива, захваљујући присуству интермедијерних филамената цитоскелета везаних за компоненте базалне ламине..
Ћелијски спојеви у биљкама
У биљном краљевству недостаје већина горе описаних ћелијских спојева, са изузетком функционалног пандана који подсећа на прорезе.
Код биљака, цитоплазме суседних ћелија повезане су путевима или каналима који се називају плазмодезми.
Ова структура ствара континуум од једне биљне ћелије до друге. Иако се структурално разликују од прореза, они имају веома сличне улоге, дозвољавајући пролаз малих јона и молекула..
Медицинска перспектива
Са становишта медицине, ћелијски синдикати су релевантно питање. Утврђено је да се мутације у генима које кодирају протеине укључене у спојеве претварају у клиничку патологију.
На пример, ако постоји одређена мутација у гену која кодира за одређени тип клаудина (један од протеина који посредује у интеракцијама у уским чвориштима), он изазива ретку болест код људи..
То је синдром губитка магнезијума у бубрезима, а симптоми укључују малу количину магнезијума и конвулзије.
Поред тога, откривено је да је мутација у гену која кодира за протеински нектин 1 одговорна за синдром расцјепа непца или расцјеп усне. Ово стање се сматра једном од најчешћих малформација код новорођенчади.
Мутације у гену нектина 1 такође су повезане са другим стањем које се зове ектодермална дисплазија и које утиче на кожу, косу, нокте и зубе људи..
Фолијарни пемфигус је булозна патологија коже одређена аутоантитијелима против десмоглеина 1, кључни елемент који је одговоран за одржавање кохезивности епидермиса.
Референце
- Албертс, Б., Браи, Д., Хопкин, К., Јохнсон, А.Д., Левис, Ј., Рафф, М., ... & Валтер, П. (2015). Основна ћелијска биологија. Гарланд Сциенце.
- Цоопер, Г. М., & Хаусман, Р. Е. (2000). Ћелија: Молекуларни приступ. Синауер Ассоциатес.
- Цуртис, Х., & Барнес, Н.С. (1994). Позив на биологију. Мацмиллан.
- Хилл, Р.В., Висе, Г.А., Андерсон, М., & Андерсон, М. (2004). Анимал пхисиологи. Синауер Ассоциатес.
- Карп, Г. (2009). Ћелијска и молекуларна биологија: концепти и експерименти. Јохн Вилеи & Сонс.
- Киерсзенбаум, А., & Трес, Л. (2016). Хистологија и ћелијска биологија: увод у патологију. Елсевиер Бразил.
- Лодисх, Х., Берк, А., Дарнелл, Ј.Е., Каисер, Ц.А., Криегер, М., Сцотт, М.П., ... & Матсудаира, П. (2008). Молекуларна ћелијска биологија. Мацмиллан.
- Воет, Д., & Воет, Ј.Г. (2006). Биоцхемистри. Ед Панамерицана Медицал.