Објави:



Карактеристике, функције, типови и инхибитори топоизомераза
Тхе топоизомеразе су типови ензима изомеразе који модификују топологију деоксирибонуклеинске киселине (ДНК), генеришући и њено одмотавање као и његово намотавање и суперковање.
Ови ензими имају специфичну улогу у ублажавању торзионог стреса у ДНК тако да се могу појавити важни процеси као што је репликација, транскрипција ДНК у гласничкој рибонуклеинској киселини (мРНА) и рекомбинација ДНК..

Енози топоизомеразе су присутни у еукариотским и прокариотским ћелијама. Њено постојање је предвиђено од стране научника Ватсона и Црицка, када су процењивали ограничења која је ДНК структура представила да омогући приступ њеним информацијама (похрањеним у нуклеотидној секвенци)..
Да би се разумеле функције топоизомераза, мора се узети у обзир да ДНК има стабилну структуру двоструке хеликса, са својим ланцима ваљаним један преко другог..
Ови линеарни ланци су састављени од 2-деоксирибозе спојене фосфодиестерским везама 5'-3 ', и азотне базе у њиховој унутрашњости, као што су степенице спиралних степеница.

Тополошко проучавање молекула ДНК показало је да они могу да прихвате неколико конформација у зависности од њихове торзионе напетости: од опуштеног стања, до различитих стања намотавања које омогућавају њихово збијање.
Молекули ДНК са различитим конформацијама називају се топоизомери. Дакле, можемо закључити да топоизомеразе И и ИИ, могу повећати или смањити торзиону напетост молекула ДНК, формирајући њихове различите топоизомере.
Међу могућим ДНК топоизомерима, најчешћа конформација је суперкоилирање, које је веома компактно. Међутим, двострука хеликса ДНК мора бити одмотана од стране топоизомераза током неколико молекуларних процеса.
Индек
- 1 Карактеристике
- 1.1 Механизам општег деловања
- 1.2 Топоизомеразе и ћелијски циклус
- 2 Функције
- 2.1 Компактно складиштење генетског материјала
- 2.2 Приступ генетским информацијама
- 2.3 Регулација експресије гена
- 2.4 Посебности топоизомеразе ИИ
- 3 Врсте топоизомераза
- 3.1-Топоизомеразе типа И
- 3.2 -Топоизомеразе типа ИИ
- 3.3 - Људски топоизомеразе
- 4 Инхибитори топоизомеразе
- 4.1-Топоизомеразе као мета за хемијски напад
- 4.2-Врсте инхибиције
- 4.3 - Инхибитори топоизомеразе
- 5 Референце
Феатурес
Механизам опште акције
Неке топоизомеразе могу да опусте само негативне ДНК супер колутове, или оба ДНК суперкалива: позитивна и негативна.
Ако се кружна дволанчана ДНК одмотава на својој уздужној оси и деси се окрет левом руком (у смеру казаљке на сату), каже се да је суперкоилован негативно. Ако је скретање у смеру казаљке на сату (супротно од казаљке на сату), оно је позитивно супер-намотано.

У основи, топоизомеразе могу:
-Олакшати пролаз низа ДНК кроз рез на супротној страни (топоизомераза тип И).
-Олакшати пролаз комплетне двоструке спирале кроз расцјеп у себи, или кроз расцјеп у другој различитој двострукој хеликси (топоизомераза типа ИИ).
Укратко, топоизомеразе дјелују кроз цијепање фосфодиестерских веза, у једној или оба ланца које чине ДНК. Затим модификовати стање намотавања нити двоструке спирале (топоизомераза И) или две двоструке спирале (топоизомераза ИИ), да би се коначно вратили на везивање или везивање крајева.
Топоизомеразе и ћелијски циклус
Иако је топоизомераза И ензим који показује већу активност током С фазе (синтеза ДНК), не сматра се да зависи од фазе ћелијског циклуса.
Док је активност топоизомеразе ИИ активнија током логаритамске фазе раста ћелија и у ћелијама брзо растућих тумора.
Функције
Промена гена који кодирају топоизомеразе је смртоносна за ћелије, показујући важност ових ензима. Међу процесима у којима учествују топоизомеразе су:
Компактно складиштење генетског материјала
Топоизомеразе олакшавају складиштење генетских информација на компактан начин, јер оне стварају ДНК намотавање и суперкоиловање, омогућавајући великој количини информација да се нађу у релативно малој количини.
Приступ генетским информацијама
Ако не би било топоизомераза и њихових јединствених карактеристика, било би немогуће приступити информацијама похрањеним у ДНК. То је зато што топоизомеразе периодично ослобађају напетост увијањем које се генерише у двострукој хеликси ДНК, током њеног одмотавања, у процесима репликације, транскрипције и рекомбинације.

Ако се напетост услед торзије која настане током ових процеса не ослободи, може произвести дефектну експресију гена, прекид кружне ДНК или хромозома, чак и производњу ћелијске смрти.
Регулација експресије гена
Конформационе промене (у тродимензионалној структури) молекула ДНК излажу одређене регионе споља, које могу да интерагују са протеинима који вежу ДНК. Ови протеини имају регулаторну функцију експресије гена (позитивну или негативну).
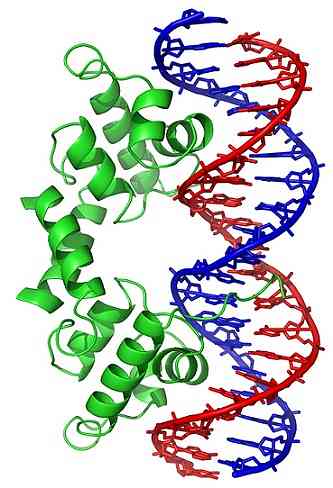
Тако, стање намотавања ДНК, генерисано дејством топоизомераза, утиче на регулацију експресије гена.
Специфичности топоизомеразе ИИ
Топоизомераза ИИ је неопходна за састављање хроматида, кондензацију и декондензацију хромозома и сегрегацију молекула кћерке ДНК током митозе.
Овај ензим је такође структурни протеин и један од главних састојака матрице ћелијског језгра током интерфазе.
Врсте топоизомераза
Постоје два главна типа топоизомераза у зависности од тога да ли су у стању да одцепе један или два ланца ДНК.
-Топоизомеразе типа И
Мономериц
Топоизомеразе типа И су мономери који ублажавају негативне и позитивне супер колуте, које настају кретањем вилице током транскрипције, и током процеса репликације и рекомбинације гена.
Топоизомеразе типа И могу се поделити на тип 1А и тип 1Б. Потоњи су они који су нађени код људи и одговорни су за опуштање супер вучне ДНК.
Тирозин на свом активном месту
Топоизомераза 1Б (Топ1Б) се састоји од 765 амино киселина подељених у 4 специфична домена. Један од ових домена има високо очуван простор који садржи активно место са тирозином (Тир7233). Све топоизомеразе на свом активном месту имају тирозин са фундаменталном улогом у целом каталитичком процесу.
Механизам дјеловања
Тирозин из активног места формира ковалентну везу са 3'-фосфатним крајем ДНК ланца, одсече је и држи везану за ензим, док пролази још један део ДНК кроз ексцизију.
Пролазак друге нити ДНК кроз исјечени ланац се постиже захваљујући конформационој трансформацији ензима, која производи отварање ДНК двоструке спирале.
Тада се топоизомераза И враћа у своју почетну конформацију и поново се веже за исечене крајеве. Ово се дешава процесом обрнутим од лома ДНК ланца, у каталитичком месту ензима. Коначно, топоизомераза ослобађа ДНК ланац.
Брзина везивања ДНК је већа од стопе ексцизије, која осигурава стабилност молекула и интегритет генома.
Укратко, топоизомераза типа И катализира:
- Изрезивање нити.
- Пролазак кроз други део кроз расцеп.
- Повезивање крајева завршава.
-Топоизомеразе типа ИИ
Димерицас
Топоизомеразе типа ИИ су димерни ензими, који раздвајају оба ланца ДНК, тако релаксирајући супершврсте које настају током транскрипције и других ћелијских процеса.
Мг Депендентс++ и АТП
Овим ензимима је потребан магнезијум (Мг++) и такође је потребна енергија која долази од разградње АТП трифосфатне везе, коју они користе захваљујући АТПази.
Два активна места са тирозином
Хумане топоизомеразе ИИ су веома сличне онима у квасцу (Саццхаромицес церевисиае), који се састоји од два мономера (подфрагменти А и Б). Сваки мономер има АТПазни домен, ау субфрагменту активни положај тирозина 782, на који се ДНК може везати. Према томе, два ланца ДНК се могу спојити са топоизомеразом ИИ.
Механизам дјеловања
Механизам дејства топоизомеразе ИИ је исти као и онај описан за топоизомеразу И, с обзиром да су два ланца ДНК расцепљена и не само један.
У активном месту топоизомеразе ИИ, стабилизован је фрагмент протеина (преко ковалентног везивања са тирозином). доубле хелик ДНК, назван "фрагмент Г". Овај фрагмент се цијепа и држи везаним за активно место ковалентним везама.
Затим, ензим дозвољава да други фрагмент ДНК, назван "Т фрагмент", прође кроз фрагмент "Г" који се цијепа, захваљујући конформацијској промјени ензима, која зависи од хидролизе АТП-а..
Топоизомераза ИИ везује два краја "фрагмента Г" и коначно опоравља своје почетно стање, ослобађајући фрагмент "Г". Затим, ДНК опушта торзиону напетост, дозвољавајући процесе репликације и транскрипције.
-Хуман топоисомерасес
Људски геном има пет топоизомераза: топ1, топ3α, топ3β (типа И); и топ2α, топ2β (типа ИИ). Најрелевантније хумане топоизомеразе су топ1 (топоизомераза типа ИБ) и 2α (топоизомераза типа ИИ).
Инхибитори топоизомеразе
-Топоизомеразе као мета за хемијски напад
Будући да су процеси катализовани топоизомеразама неопходни за преживљавање ћелија, ови ензими су добри циљеви напада да утичу на малигне ћелије. За ово, топоизомеразе се сматрају важним у лечењу многих хуманих обољења.
Лијекови који су у интеракцији с топоизомеразама тренутно су широко проучавани као хемотерапијске супстанце против ћелија рака (у различитим органима тела) и патогених микроорганизама..
-Врсте инхибиције
Лекови који инхибирају активност топоизомеразе могу:
- Унос у ДНК.
- Утицај на ензим топоизомеразе.
- Преплетање са молекулом близу активног места ензима док је ДНК-топоизомеразни комплекс стабилизован.
Стабилизација пролазног комплекса који се формира везивањем ДНК за тирозин каталитичког места ензима спречава уједињење исечених фрагмената, што може довести до смрти ћелије..
-Инхибитори топоизомераза
Међу једињењима која инхибирају топоизомеразе су следећа.
Антитуморни антибиотици
Антибиотици се користе против рака, јер спречавају раст туморских ћелија, обично ометањем њиховог ДНК. Често се називају антинеопластичним антибиотицима (против рака). Актиномицин Д, на пример, утиче на топоизомеразу ИИ и користи се код Вилмсових тумора код деце и код рабдомиосаркома..
Антхрацицлинес
Антрациклини су, међу антибиотицима, најефикаснији антиканцерогени лекови и најшири спектар. Користе се у лечењу рака плућа, јајника, материце, желуца, бешике, дојке, леукемије и лимфома. Познато је да утиче на топоизомеразу ИИ интеркалацијом у ДНК.
Први антрациклин изолован из актинобактерије (Стрептомицес пеуцетиус) је био даунорубицин. Након тога, у лабораторији је синтетисан доксорубицин, а сада се користе и епирубицин и идарубицин..
Антхракуинонес
Антракинони или антраценедиони су једињења изведена из антрацена, слична антрациклинима, који утичу на активност топоизомеразе ИИ интеркалацијом у ДНК. Користе се за метастатски рак дојке, не-Ходгкинов лимфом (НХЛ) и леукемију.
Ови лекови су пронађени у пигментима неких инсеката, биљака (франгула, сенна, рабарбара), лишајева и гљивица; као иу хоелити, која је природни минерал. У зависности од њихове дозе, оне могу бити канцерогене.
Међу овим једињењима је митоксантрон и његов аналог лосоксантрон. Они спречавају пролиферацију малигних ћелија тумора, иреверзибилно се вежу за ДНК.
Епидофилотоксини
Подофилотоксини, као што су епидофилотоксини (ВП-16) и тенипозид (ВМ-26), формирају комплекс са топоизомеразом ИИ. Користе се против рака плућа, тестиса, леукемије, лимфома, рака јајника, карцинома дојке и малигних интракранијалних тумора, између осталих. Они су изоловани од биљака Подопхиллум нотатум и П. пелтатум.
Аналози камптотецина
Кампотецини су једињења која инхибирају топоизомеразу И, а међу њима су иринотекан, топотекан и дифломотекан.
Ова једињења су коришћена против рака дебелог црева, плућа и дојке, и добијена су природно од коре и лишћа дрвених врста. Цамптотхеца ацумината кинеске брескве и Тибет.
Природна инхибиција
Структурне промјене топоизомераза И и ИИ могу се појавити потпуно природно. То се може десити током неких догађаја који утичу на његов каталитички процес.
Међу овим промјенама, можемо споменути формирање пиримидинских димера, неусклађености азотних база и других догађаја узрокованих оксидативним стресом..
Референце
- Андерсон, Х., & Роберге, М. (1992). ДНА топоизомераза ИИ: преглед њеног учешћа у структури хромозома, репликацији ДНК, транскрипцији и митози. Целл Биологи Интернатионал Репортс, 16 (8): 717-724. дои: 10.1016 / с0309-1651 (05) 80016-5
- Цххатривала, Х., Јафри, Н., & Салгиа, Р. (2006). Преглед инхибиције топоизомеразе код рака плућа. Цанцер Биологи & Тхерапи, 5 (12): 1600-1607. дои: 10.4161 / цбт.5.12.3546
- Хо, И.-П., Ау-Иеунг, С.Ц.Ф., & То, К.К.В. (2003). Антиканцерни агенси на бази платине: иновативне стратегије дизајна и биолошке перспективе. Медицинал Ресеарцх Ревиевс, 23 (5): 633-655. дои: 10.1002 / мед.10038
- Ли, Т.-К., & Лиу, Л.Ф. (2001). Смрт туморских ћелија изазвана лековима који циљају на топоизомеразу. Годишњи преглед фармакологије и токсикологије, 41 (1): 53-77. дои: 10.1146 / аннурев.пхармток.41.1.53
- Лиу, Л.Ф. (1994). ДНА Топоисомерасес: Топоисомерасе-Таргетинг Другс. Ацадемиц Пресс. пп 307
- Осхерофф, Н. и Бјорнсти, М. (2001). ДНА Топоисомерасе. Ензимологија и дроге. Вол. Хумана Пресс. пп 329.
- Ротхенберг, М. Л. (1997). Инхибитори топоизомеразе И: Преглед и ажурирање. Анналс оф Онцологи, 8 (9), 837-855. дои: 10.1023 / а: 1008270717294
- Риан Б. (2009, децембар 14). Топоизомераза 1 и 2. [Видео фајл]. Преузето са иоутубе.цом