Објави:



Карактеристике Фораминифера, морфологија, класификација, животни циклус
Тхе фораминифера они су група амоебоид протозоа, неке морске и друге слатке воде. Појавили су се на почетку примарне ере (камбрија), а њихови насљедници су населили данашње океане. Могу се наћи из обалних подручја (хипо или хиперсалина) до дна океана, а од тропа до хладног арктичког и антарктичког океана.
Његова дистрибуција зависи од неколико фактора. На пример, постоје врсте које подржавају велике и честе промене температуре, док друге не могу да преживе, тако да термичка структура океана означава важне разлике између асоцијација фораминифера.

Исто тако, дубина је такође одлучујући фактор у дистрибуцији фораминифера због њихове директне учесталости у продору светлости. С друге стране, притисак је повезан са претходним факторима (температура и дубина), директно утичући на растворљивост ЦО2, што утиче на излучивање калцијум карбоната за формирање шкољки.
С друге стране, енергија воде у плитким подручјима је релевантан елемент јер утиче на врсту супстрата (тврда или мека) и дистрибуцију нутријената.
Исто тако, други фактори као што су салинитет, замућеност воде, пХ, присуство елемената у траговима и / или органских компоненти, струје, брзина седиментације, могу одредити, локално, дистрибуцију фораминифера.
Индек
- 1 Познате врсте
- 2 Карактеристике
- 2.1 Фактори укључени у величину и морфологију фораминифера
- 3 Таксономија
- 4 Класификација
- 4.1 Атхаламеа
- 4.2 Монотхаламеа
- 4.3 Ксенопхиопхореа
- 4.4 Туботхаламеа
- 4.5 Глоботхаламеа
- 5 Морфологија
- 5.1-Величина
- 5.2 -Протоплазма
- 5.3 - Скелет или љуска
- 5.4 - Врсте Фораминифера
- 5.5 -Сеудоподи
- 6 Животни циклус
- 7 Репродукција
- 8 Исхрана
- 9 Апликације
- 9.1 Случај Маурицијуса
- 10 Референце
Познате врсте
Тренутно је познато више од 10.000 врста, а око 40.000 је изумрло. Неке врсте имају као подморје морско дно, то јест, они су бентички организми, често живе камуфлирани на пијеску који чини дио епифауне (епибетоницос) или могу живјети под пијеском (ендобетоницос). Због тога су познати и као живи пијесак.
Они могу да живе и на биљкама, у којима се одмарају као епифити, па чак и многи од њих бирају живот у сесилу, тј..
Док други фораминифери живе плутајући на различитим дубинама у океану (између 0 и 300 м), тј. Они су планктонски животни део морског микропанктона. Ови облици су оскуднији и мање разнолики.
Највећи и најсложенији планктонски фораминифери су чешћи у тропским и субтропским срединама. Док су у високим географским ширинама ови организми обично ретки, мањи и на врло једноставне начине.
Феатурес
Карактеристика која се истиче у фораминифери је скелет или љуска, структура која је омогућила да се изумрли облици могу проучавати у облику морских микрофосила, који се таложе на дну мора..
Дакле, љуска је основни елемент за разликовање фораминифера и једина је структура организма која фосилизира. Ови фосили су веома обилни у морском седименту, такође учествују у формирању седиментних стена.
Главна хемијска једињења шкољки су калцит, арагонит и силицијум диоксид. Облик и димензија ембрионалне коморе зависи од њеног порекла, било да је то производ сексуалне или асексуалне репродукције.
За време њихове онтогенезе, фораминифера контролише раст и величину комора. Ова контрола се врши кроз дужину и распоред псеудоподијалних струја, јер су псеудоподи одговорни за формирање органске мембране која претходи минералној љусци..
Овај процес је веома важан за одржавање ћелијских процеса, пошто камера делује као биореактор.
Фактори који интервенишу у величини и морфологији фораминифера
Треба напоменути да величина и коначна морфологија коју фораминифера може да усвоји зависи од неколико фактора, укључујући:
- Облик и димензије ембрионалне коморе.
- Број стадијума раста до стадијума одраслих (тј. Број комора фораминифера).
- Облик камере и њене модификације током онтогенезе.
- Распоред камера.
Већи фораминифери поседују стратешке обрасце раста како би одржали волумен константних комора без прекорачења оптималне величине. Ове стратегије се састоје од подјеле камера у неколико одјељака званих клике.
Ови кластери су распоређени на такав начин да осигуравају транспортне и регулаторне функције између протоплазме унутар комора и извана. То јест, све камере и клике су савршено међусобно повезане.
Распоред комора може пратити правоцртну или спиралу. То ће зависити од положаја псеудоподијалних струја и локације отвора или отвора у комори.
Такономи
Домаин: Еукариа
Кингдом: Протиста
Без домета: САР Супергроуп
Суперпхилум: Рхизариа
Тип: Фораминифера
Класе и наредбе
- Атхаламеа (Ретицуломикида)
- Монотхаламеа (Аллогромиида, Астрорхизида, Комокиида)
- Ксенопхиопхореа (Псамминиде, Станномид)
- Туботхаламеа(Фусулинида, Инволутинида, Милиолиде, Силицолоцулиниде, Спириллиниде)
- Глоботхаламеа(Литуолида, Лофтусиида, Сцхлумбергеринида, Тектулариида, Троцхаминминида, Роталиде, Булиминиде, Глобигериниде, Робертинида, Цартериниде, Лагенида).
Класификација
Иако још има много ствари које треба разјаснити, до сада можемо разликовати 5 класа:
Атхаламеа
Овде можете пронаћи фораминифере који немају шкољку или који су голи.
Монотхаламеа
Укључује бетонски фораминифер који представља органску или аглутинирану љуску са једном комором.
Ксенопхиопхореа
У овом случају, фораминифера је специјализованог Бетонског типа велике величине, мултинуклеарне и аглутиниране љуске. Обично су детритивори или сапрофаги, тј. Добијају храну од детритуса или органске материје у разградњи.
Туботхаламеа
Ово укључује бетонске фораминифере, које имају вишеструке тубуларне коморе, бар у јувенилној фази, које могу бити намотане у спиралу, са везаном или калцантном љуском..
Глоботхаламеа
Ова класификација укључује и бетонски и планктонски фораминифера са мултикамералним шкољкама глобуларне, аглутиниране или карбонатне форме. Шкољке могу бити унисерате, бисериада, трисериада или троцоспираладас.
Међутим, ова класификација је у сталној еволуцији.
Морфологија
-Величина
Величина фораминифера се обично креће између 0,1 и 0,5 цм, при чему неке врсте могу да мере од 100 μм до 20 цм..
-Протоплазма
Фораминифера се формира протоплазматском масом која чини ћелију фораминифера.
Протоплазма је обично безбојна, али понекад може да садржи мале количине органских пигмената, липидног материјала, симбиотских алги или једињења гвожђа који дају боју.
Протоплазма се састоји од унутрашњег дијела који се назива ендоплазма и вањске ектоплазме.
У ендоплазми је заштићена љуском иу њој се органеле дистрибуирају као дигестивне вакуоле, језгра, митохондрије, грануле, Голгијеви апарати или рибозоми. Из тог разлога се понекад назива грануларна ендоплазма. Ектоплазма је прозирна и одатле одлазе помични псеудоподи.
Протоплазма је ограничена споља органском мембраном коју чине слојеви мукополисахарида.
Протоплазматска маса се шири из љуске кроз један или више отвора (пора) и покрива споља (екстра камерална протоплазма), и тако се формирају псеудоподије..
-Скелет или љуска
Фораминифера фиксно фиксира своју целуларну површину, конструкцијом минералног скелета (љуске).
Шкољку чине коморе раздвојене септама, али истовремено комуницирају једна са другом кроз рупе за међусобно повезивање које се називају форамина, отуда и име фораминифера. Хемијски састав скелета или љуске чини их структурама које се врло лако фосилизирају.
Унутрашњост комора је прекривена органским материјалом врло сличним хитину. Поред тога, љуска може имати главне отворе; Такође можете да поседујете поре или да их не поседујете.
Минерална шкољка може бити формирана од једног комора (примитивног фораминифера или моноталамоса), или коморе, која расте континуирано или неколико комора које се формирају у узастопним фазама, у компликованом систему дисконтинуираног раста (фораминифера полиламас)..
Овај последњи процес се састоји у додавању раније формиране љуске, а на стратешким местима, новог скелетног материјала.
Многе фораминифере могу да изаберу материјал за формирање своје љуске према њиховом хемијском саставу, величини или облику, јер су маргиналне псеудоподијске струје које су у контакту са супстратом у стању да га препознају..
-Врсте Фораминифера
Према облику конструкције љуске, могу се сврстати у три главне врсте Фораминифера:
Аглутинисани (или зрнасти)
У овој врсти љуске, фораминифера сакупља са својим псеудоподама велику количину органских материјала доступних у околини у којој живе, која затим аглутинира, као што су минерална зрна, спикуле спужви, дијатомеје итд..
Већина аглутинисаних фораминифера цементира своју љуску калцијум карбонатом, али ако ово једињење није присутно у околини, као на пример они који живе у дубоким деловима океана где нема калцијума, то могу да ураде са силикатним, ферругинозним, органским цементима, итд.
Порцелаин
У овом случају, љуска се формира преко иглица магнезијумовог калцита које се синтетизују у Голгијевом апарату фораминифера.
Ове игле се транспортују и акумулирају са спољне стране и могу служити као елементи за спајање страних структура (Цемент) или директно из вањског костура. Налазе се у хиперсалиној средини (> 35% салинитета).
Обично су незупчане, тј. Обично имају псеудо поре које не пролазе кроз љуску у потпуности.
Хиалинес
Оне се формирају растом кристала калцита захваљујући органском шаблону, који се формира процесом који се зове биоминерализација (минерализација ин ситу), који се изводи споља у протоплазматско тело.
Одликује их транспарентност, због танкости зида. Они су такође перфорирани где је локација, густина и пречник пора променљива у зависности од врсте.
-Псеудоподс
Ова структура се користи за мобилизацију, везивање за супстрате, хватање плена и стварање скелета. За ретракцију и проширење псеудопода фораминифера има софистицирану мрежу микротубула распоређених у више или мање паралелних редова.
Продужетак псеудопода може достићи два или три пута дужину тела и може чак бити и до 20 пута дужине. То ће зависити од сваке поједине врсте.
Тип кретања током померања је директно повезан са обликом љуске и положајем отвора (одакле потичу псеудоподи)..
Али већина фораминифера се креће на следећи начин: псеудоподи се везују за супстрат, а затим гурају остатак ћелије. Кретање на овај начин може напредовати брзином од приближно 1 до 2,5 цм / сат.
С друге стране, псеудоподи фораминифера се називају Гранурретицулоподиа, јер у унутрашњости псеудопода постоји двосмјерни цитоплазматски проток који носи грануле.
Грануле се могу састојати од честица различитих материјала, митохондрија, дигестивних или отпадних вакуола, синбиотских динофлагелата, итд. Из тог разлога један од синонима групе је Гранулоретицулоса.
Још једна битна карактеристика псеудопода је да имају тенденцију да буду дуги, танки, разгранати и веома обилни, формирајући на тај начин мрежу ретицулоподије стогом (анастомоза)..
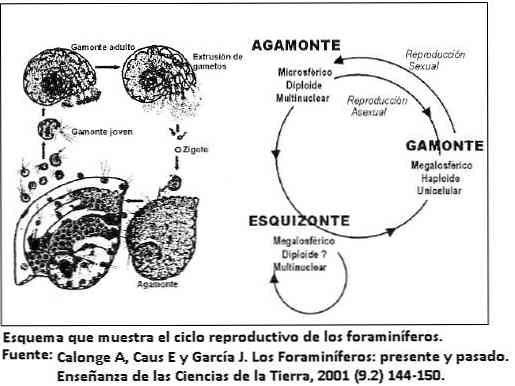
Животни циклус
Животни циклус фораминифера је обично кратак, обично неколико дана или недеља, али у великим облицима животни циклус може достићи две године..
Трајање ће зависити од животне стратегије коју фораминифера прихвата. На пример, мали облици са једноставном морфологијом развијају кратку опортунистичку стратегију.
Док велике форме и са изузетно сложеном морфологијом љуске развијају конзервативну животну стратегију.
Ово последње понашање је веома неуобичајено у једноћелијским организмима; омогућава им да одрже јединствену густину насељености и споро расту.
Репродукција
Већина фораминифера има две морфологије, са генерационом изменом у складу са типом репродукције, сексуалним или асексуалним, осим планктонских фораминифера које се репродукују само сексуално..
Ова промена у морфологији се назива диморфизам. Настали облик сексуалне репродукције (гамогониа) назива се гамон, док асексуална репродукција (шизогонија) даје облик шизонта. Оба су морфолошки различита.
Неки фораминифери координирају циклус репродукције са сезонским циклусом, како би оптимизирали кориштење ресурса. Није неуобичајено видети неколико континуираних асексуалних репродукција пре него што се сексуална генерација јави у бетонским формама.
То објашњава зашто су облици шизонта обилнији од оних гамонта. Гамон у почетку представља једно једро, а затим се дели да производи бројне гамете.
Док је шизонт мултинуклеарни и након мејоза фрагменти формирају нове гамете.
Нутритион
Фораминифера је карактеристична по томе што је хетеротрофна, тј. Хране се органском материјом.
У овом случају фораминифера се храни углавном дијатомејама или бактеријама, али се друге веће врсте хране нематодама и раковима. Затвореници су заробљени кроз псеудоподе.
Такође, ови организми могу да користе симбионтске алге различитих типова, као што су зелене, црвене и златне алге, као и дијатомеје и динофлагелате, а чак може да постоји и веома сложена разноликост многих од њих у истој особи..
С друге стране, неке врсте фораминифера су клептопластичне, што значи да хлоропласти из унесених алги постају део фораминифера да би наставили да обављају функцију фотосинтезе..
Ово представља алтернативни начин производње енергије за живот.
Апплицатионс
Богатство фосилног записа фораминифера дуж геолошког времена, еволуције, сложености и величине чини их омиљеним инструментом за проучавање садашњости и прошлости Земље (геолошки сат).
Због тога је његова велика разноликост врста веома корисна у истраживањима биостратиграфског, палеоеколошког, палеоцеанског типа..
Међутим, она такође може помоћи да се спрече еколошке катастрофе које могу да утичу на економију, пошто промене у популацији фораминифера указују на промене у окружењу.
На пример, фораминифера без шкољки, осетљиви су на промену животне средине и брзо реагују на промене окружења које их окружује. Због тога су идеалне индикаторске врсте за проучавање квалитета и здравља гребенасте воде.
Случај Маурицијуса
Такође, неки догађаји су нас навели на размишљање о овоме. Такав је случај са феноменом који је примећен на Маурицијусу, где је део белог песка на плажи нестао и сада га морају увозити са Мадагаскара да би одржали туристички ток.
И шта се тамо десило? Одакле долази пијесак? Зашто је нестао??
Одговор је следећи:
Песак није ништа друго до акумулација калцијум карбонатних шкољки многих организама, међу њима и фораминифера које се вуку до руба плаже. Нестанак песка узрокован је прогресивним и континуираним смањењем производње карбоната.
То се десило као посљедица загађења мора душиком и фосфором који допиру до обала због прекомјерне употребе гнојива у сјетви одређених производа као што је шећерна трска..
Због тога је важно проучавати фораминифере у друштвеним наукама, како би се спречиле еколошке катастрофе, као што је горе описано, које директно утичу на привреду и друштво..
Референце
- Википедиа цонтрибуторс. Фораминифера [онлине] Википедиа, Слободна енциклопедија, 2018 [датум консултација: 1. новембар 2018]. Доступно на ес.википедиа.орг.
- Цалонге А, Цаус Е и Гарциа Ј. Фораминифера: садашњост и прошлост. Настава наука о Земљи, 2001 (9.2) 144-150.
- Хромиц Т. Биодиверзитет и екологија микробентоса (Фораминифера: Протозоа), између Боца дел Гуафо и Голфо де Пенас (43º-46º с), Чиле. Циенц. Тецнол. 30 (1): 89-103, 2007
- Хумпхреис АФ, Халфар Ј, Ингле ЈЦ, ет ал. Утицај температуре мора, пХ и хранљивих састојака на расподелу и карактеристику плитког бентосног фораминифера у малом обиљу у Галапагосу. ПЛоС Оне 2018; 13 (9): е0202746. Објављено 2018 Сеп 12. дои: 10.1371 / јоурнал.поне.0202746
- Де Варгас Ц, Норрис Р, Занинетти Л, Гибб СВ, Павловски Ј. Молекуларни докази криптичне специације у планктонским фораминиферима и њихов однос према оцеанским провинцијама. Проц Натл Ацад Сци У С А. 1999; 96 (6): 2864-8.